Mechanizm, w jakim niekiedy z pozoru nieefektywne zabiegi osteopatyczne przekazywane poprzez dźwignię narządu ruchu czy szerzej – powięziową sferę ścienną (parietalną) osiągają zwielokrotniony efekt w postaci regulacji homeostazy, jest nadal przedmiotem żywiołowej dyskusji.
Sceptycy krytykują wygórowane ich zdaniem aspiracje terapii manualnej i z trudnością akceptują niektóre spektakularne (szybkie i powtarzalne) jej efekty, jak choćby uwolnienie kolki niemowlęcej po terapii osteopatycznej podstawy czaszki i punktów wyjścia nerwu błędnego. Dla entuzjastów efekty terapii manualnej są przedmiotem dumy z „ponadnaturalnych” zdolności, ale niejednokrotnie stają się pułapką, gdy w sposób trudny do zaakceptowania dla środowiska lekarskiego próbują „leczyć wszystko” i tworzą niechcący medycynę alternatywną. Wydaje się, że u podstaw owych tajemniczych efektów terapii osteopatycznej leżą procesy autoregulacji zachodzące za pośrednictwem autonomicznego układu nerwowego (AUN).
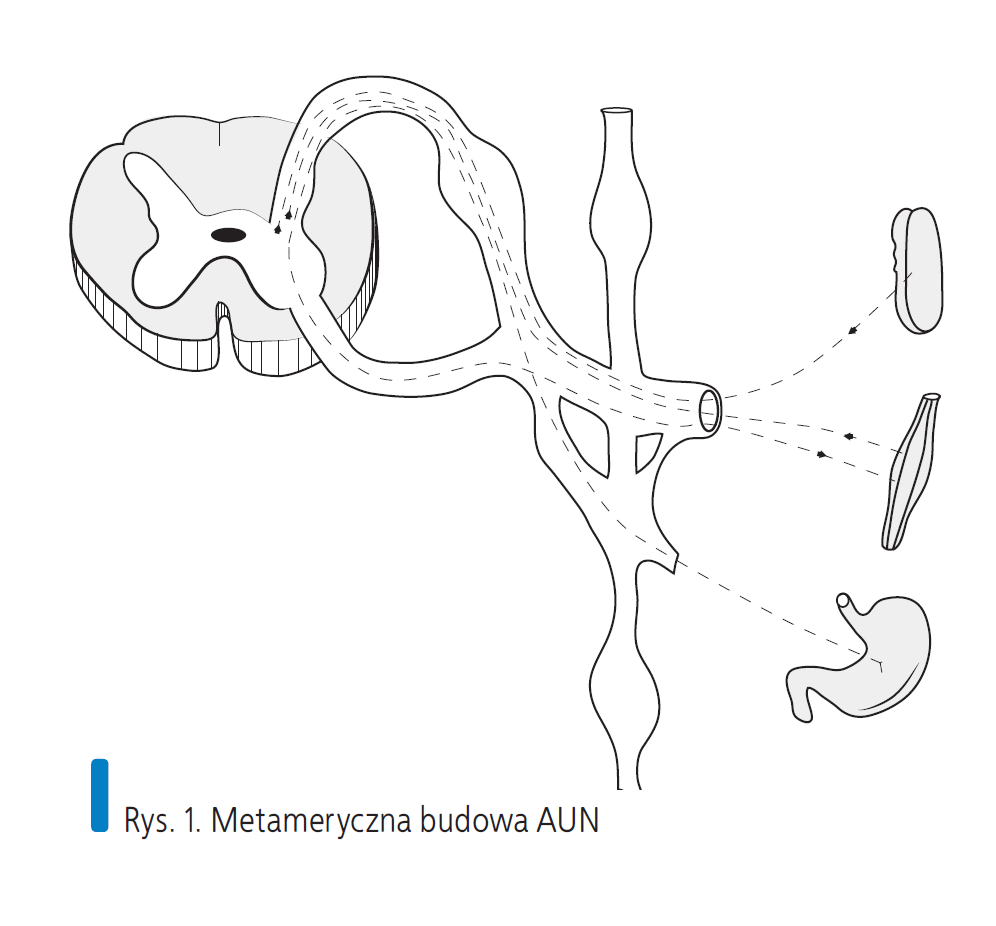
Rys. 1. Metameryczna budowa AUN
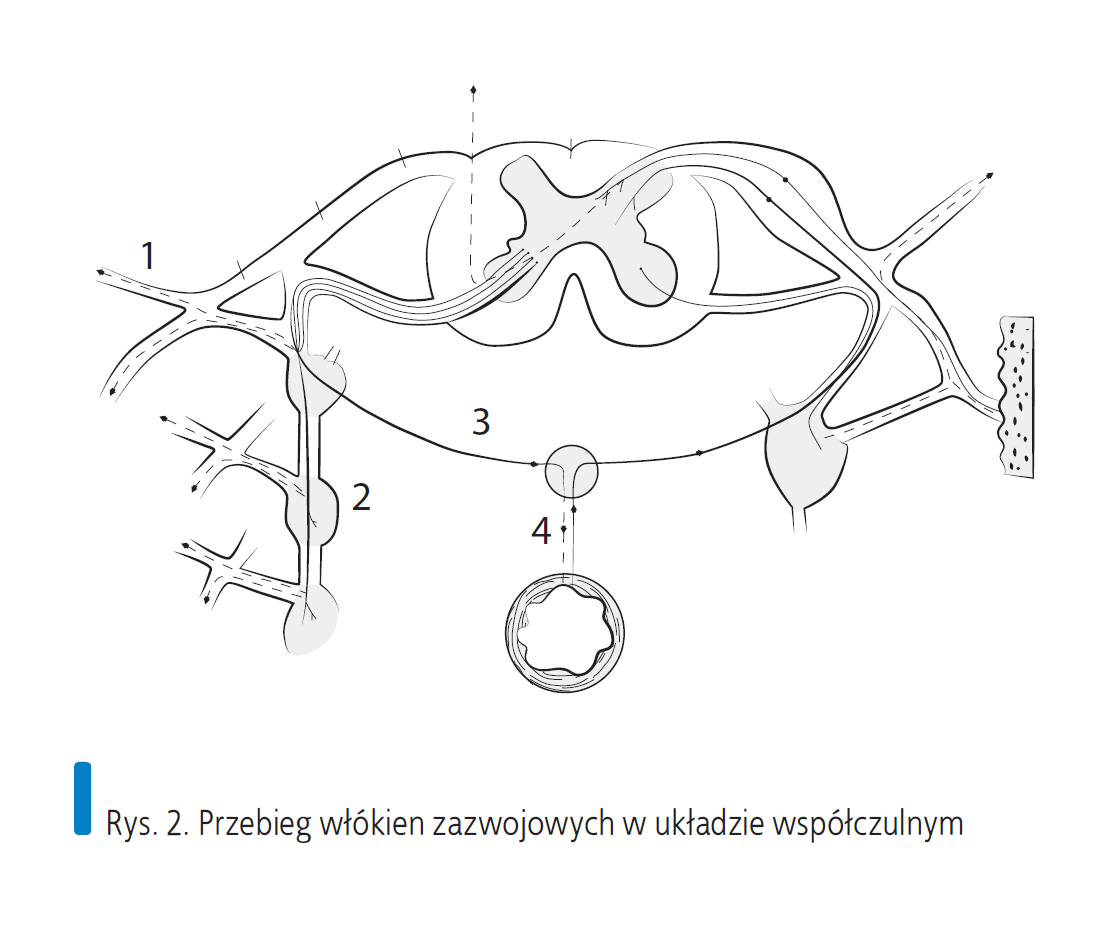
Rys. 2. Przebieg włókien zazwojowych w układzie współczulnym
Osteopatia
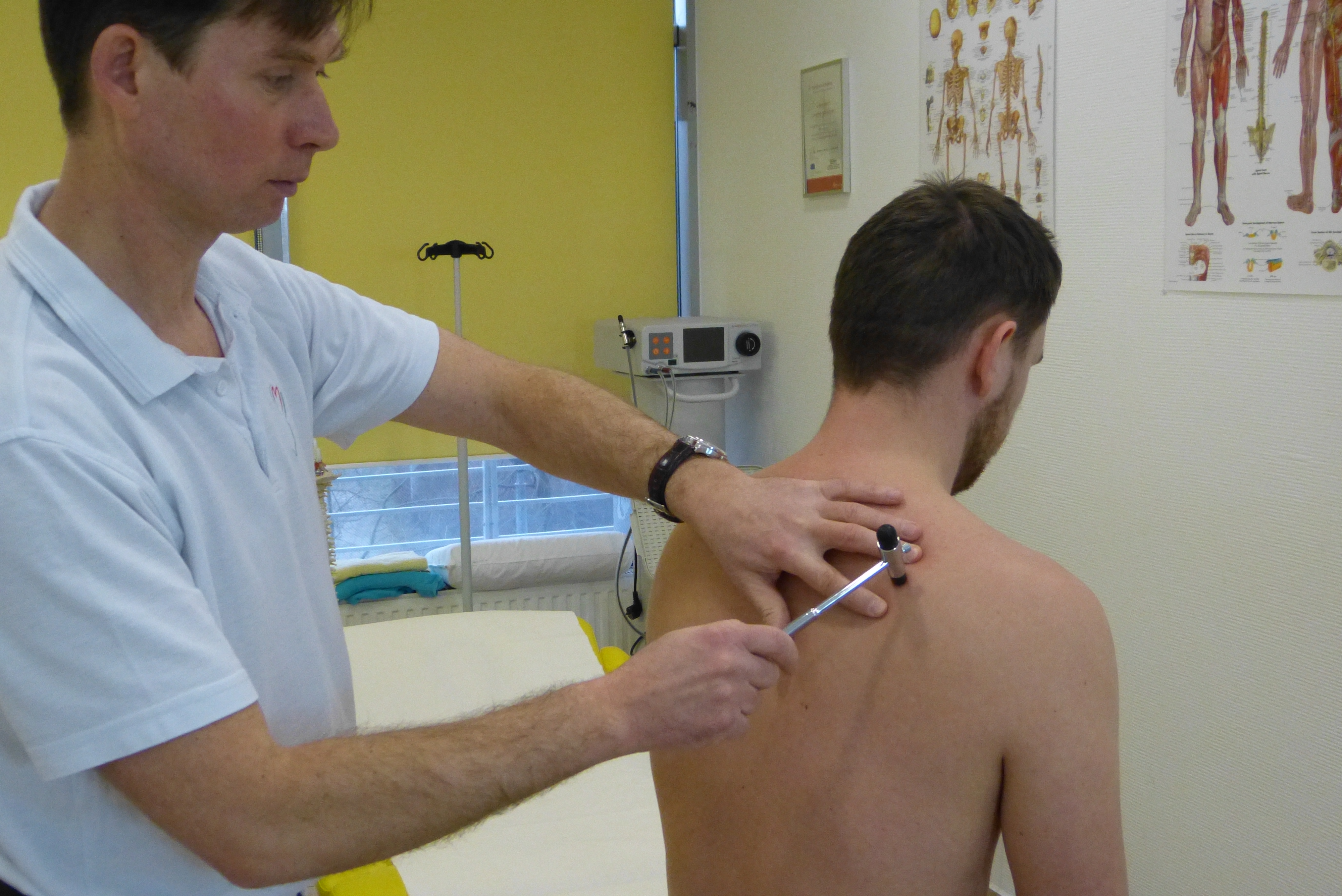
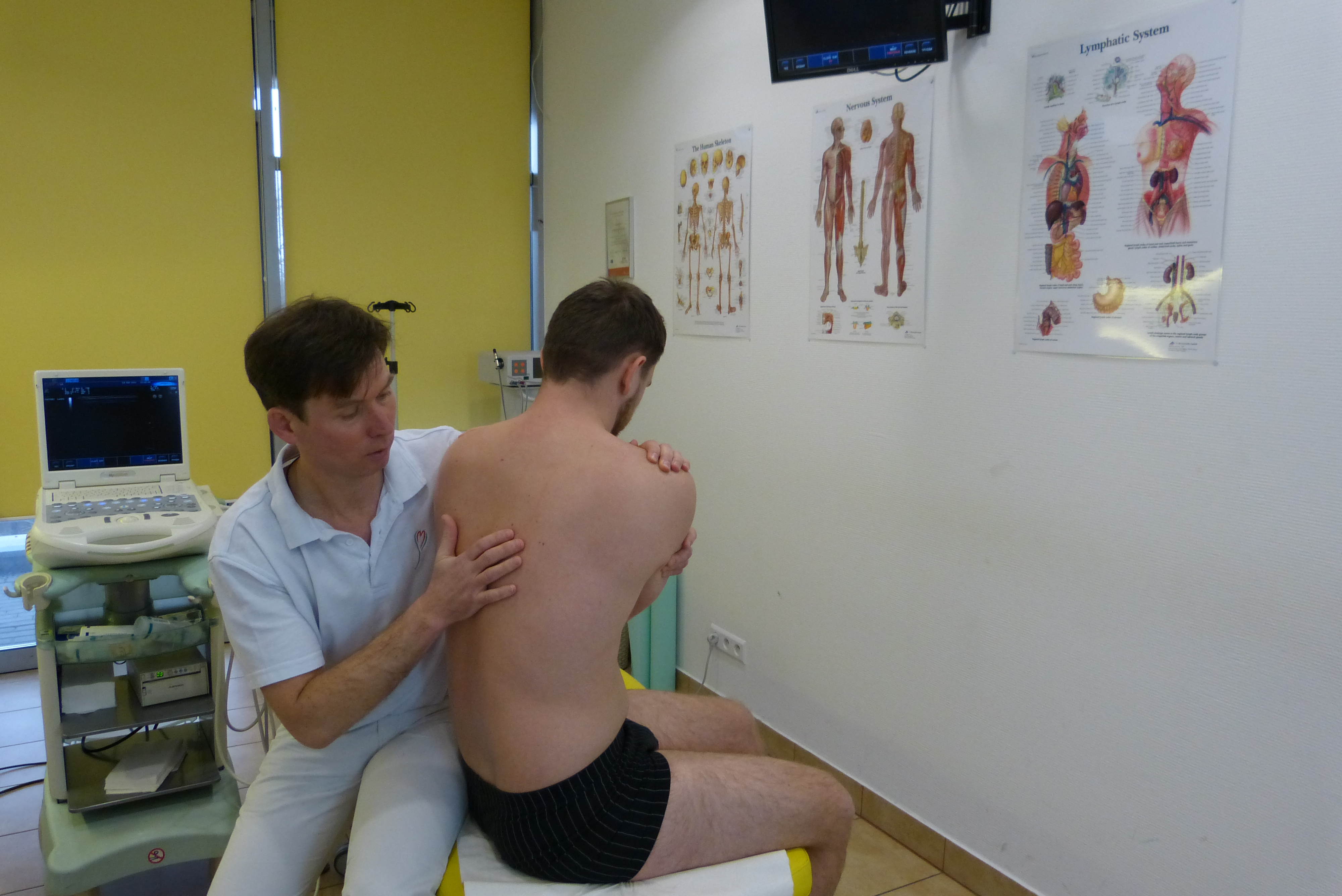
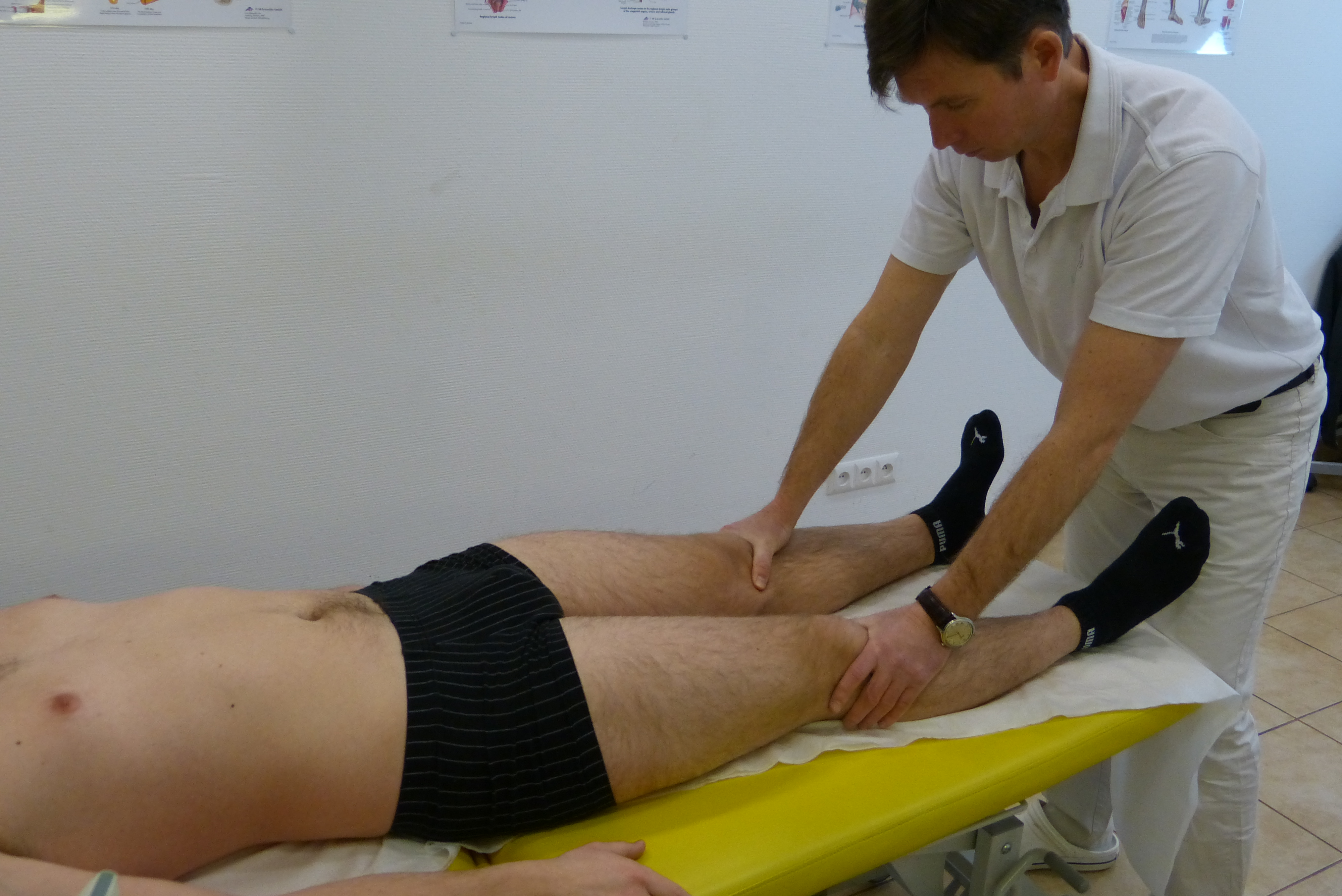
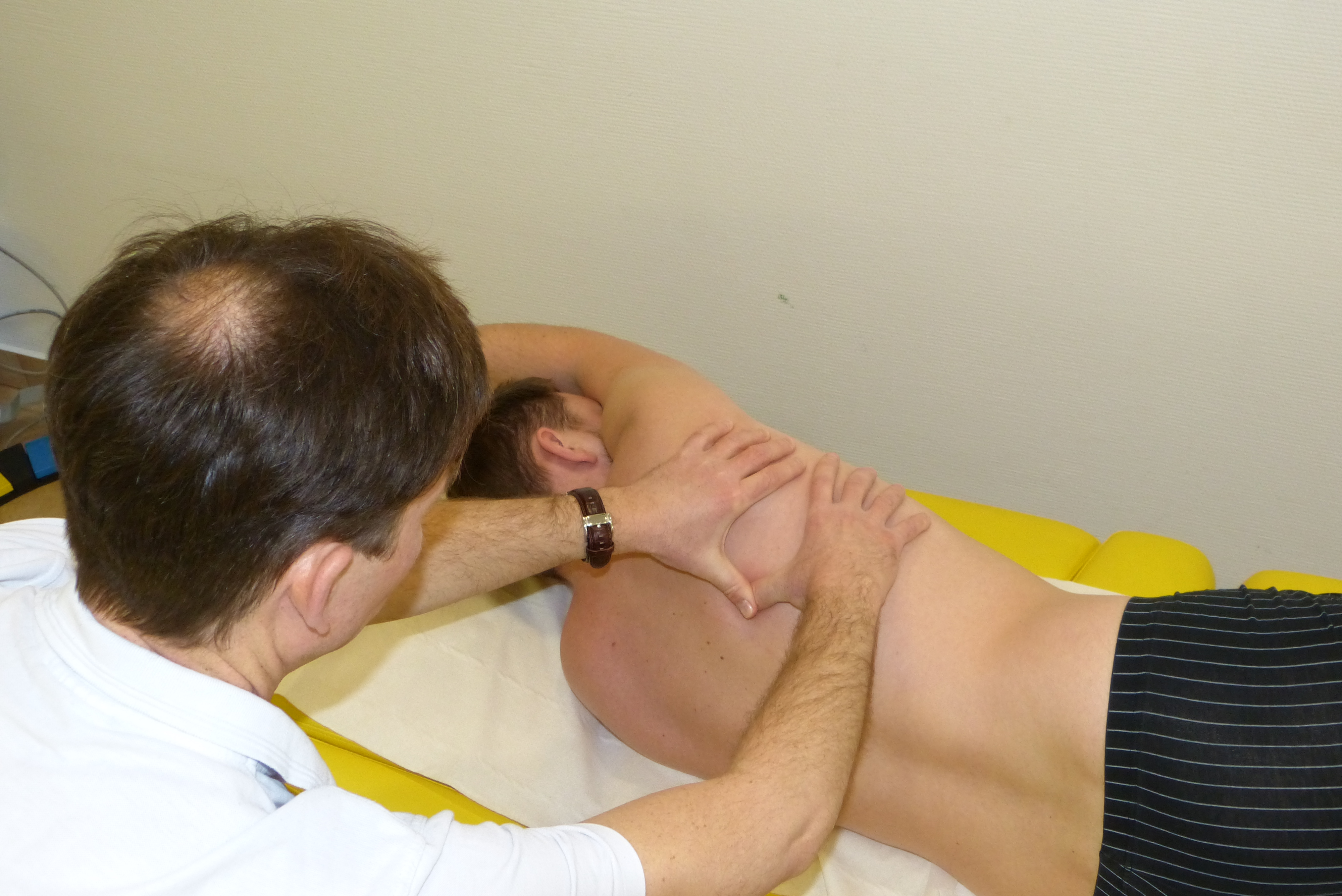
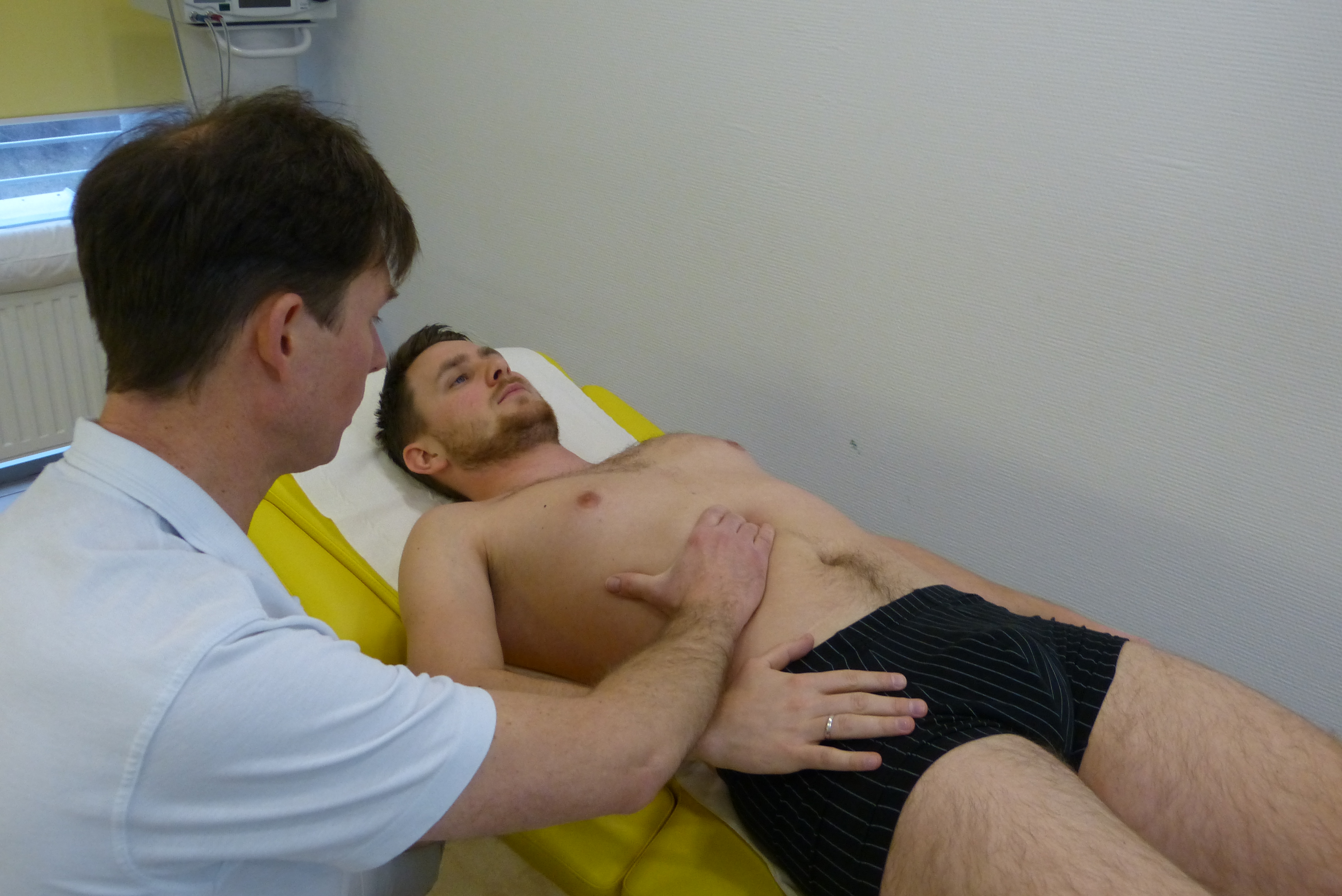
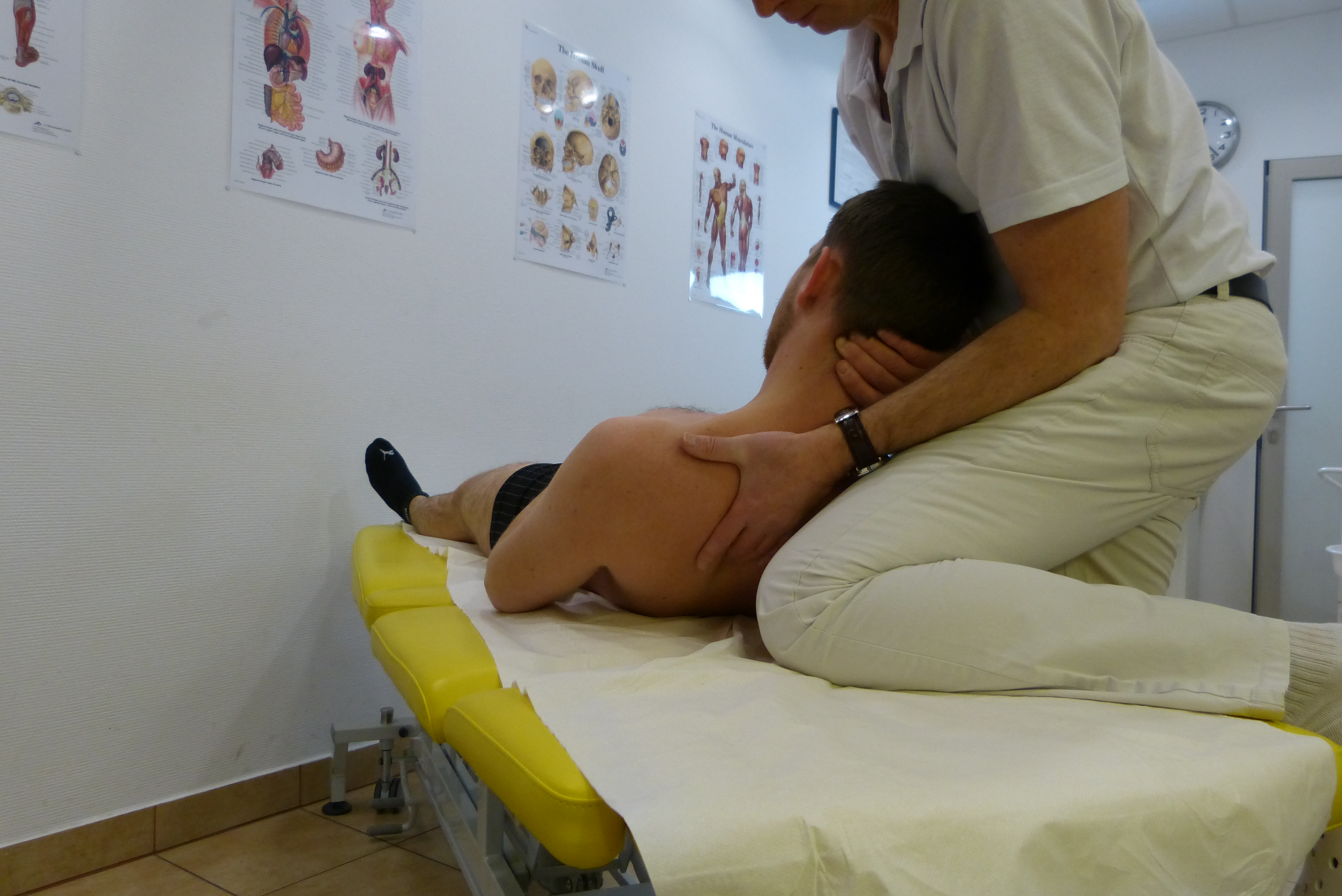
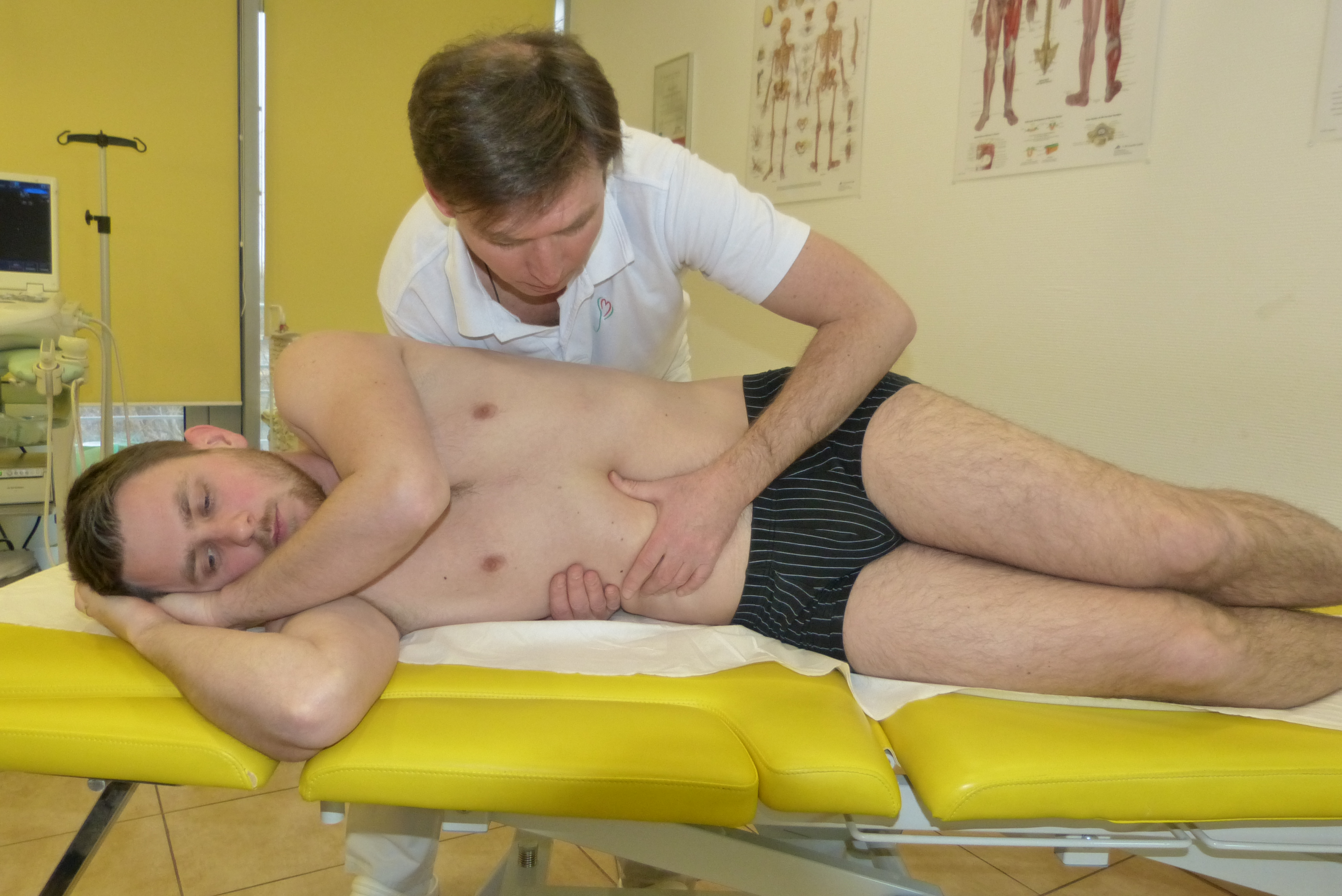
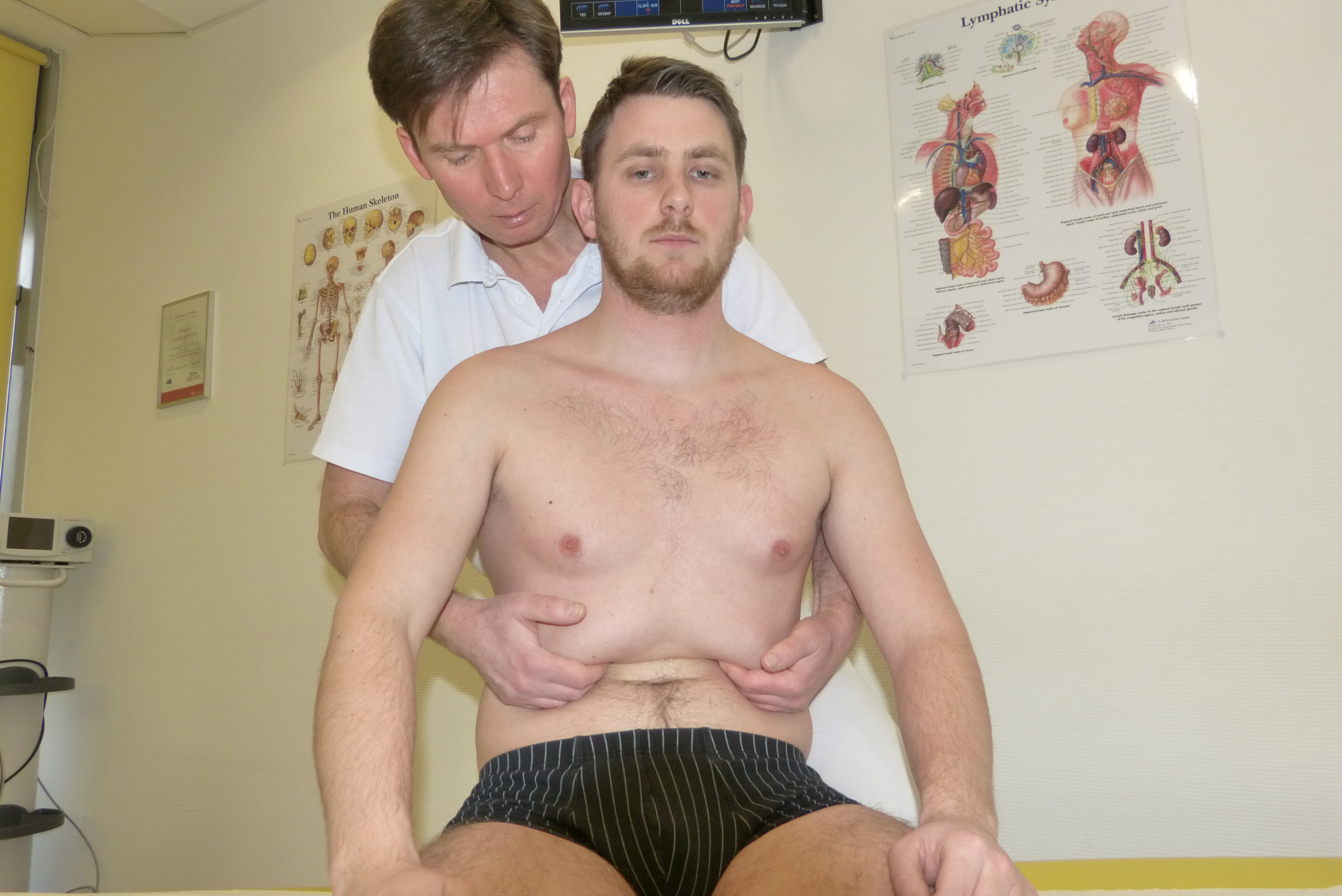
Anatomia i fizjologia
Autonomiczny układ nerwowy stanowi obok centralnego i obwodowego układu nerwowego trzeci (bodaj najbardziej fascynujący ze względu na zasięg, budowę i zakres oddziaływania) obszar unerwienia. Dociera do każdego narządu w organizmie, moduluje funkcje czuciowe, ruchowe i wydzielnicze, uczestniczy w procesach gojenia, wzrostu, ale też w procesach poznawczych i behawioralnych. O jego odrębności decyduje również to, że transmisja synaptyczna w układzie autonomicznym ma charakter chemiczny (acetylocholina i noradrenalina), a nie elektryczny [1]. Działa on niezależnie (autonomicznie) i w sposób ciągły, bez świadomego wysiłku jego „użytkownika”, ale oczywiście nie bez kontroli ośrodków wyższych, takich jak układ limbiczny, podwzgórze i twór siatkowaty. Jego części znajdują w takich obszarach mózgu, jak kora przedniej obręczy, wyspa, ciało migdałowate, hipokamp. Jest obecny w rdzeniu przedłużonym (jądra nerwów czaszkowych), rdzeniu kręgowym i zwojach przykręgowych pnia współczulnego, gdzie tworzy jedyne w swoim rodzaju nagromadzenie komórek nerwowych poza centralnym układem nerwowym. Jego efektory rozsiane są w skórze i jej gruczołach, w mięśniach gładkich przewodu pokarmowego, układu oddechowego, moczowo-płciowego i komórkach wydzielniczych gruczołów dokrewnych (np. rdzeń nadnerczy). Zawiera włókna dośrodkowe (trzewno-czuciowe), którymi wysyła informację do rdzenia i mózgu o stanie narządów wewnętrznych oraz włókna odśrodkowe (trzewno-ruchowe) drogą, którą wysyła impulsy do efektorów – mięśni gładkich, mięśnia sercowego, skóry, gruczołów itp. [2].
Autonomiczny układ nerwowy rozwija się w sposób metameryczny, znajdując swoją reprezentację rdzeniową w odpowiadającym mu obszarach skóry, naczyń, mięśniówki gładkiej czy torebkach narządów trzewnych zaopatrywanych przez dany nerw rdzeniowy.
Dzieje się tak, ponieważ układ autonomiczny w pewnym stopniu korzysta z „gościnności” układu somatycznego. Drogą nerwów obwodowych biegną włókna układu autonomicznego odśrodkowe i dośrodkowe, wychodząc z rdzenia poprzez korzenie, a po przełączeniu w zwojach wracają do swoich somatycznych nerwów rdzeniowych. Pozostałe włókna korzystają z rusztowania naczyń, oplatając je jak siatka i nosząc nazwy pochodzące od naczynia, na którym spoczywają (t. kręgowa, t. podobojczykowa, aorta, tętnice nerkowe, biodrowe i inne), inne wtapiają się w powięzi (krezki jelita, więzadła mocujące trzewia), biegną torem nerwów czaszkowych (III, VII, IX i X) lub tworzą własne struktury, takie jak zwoje, sploty, pnie i nerwy trzewne.
Ciała neuronów układu współczulnego wysyłające włókna odśrodkowe (trzewno-ruchowe) usytuowane są w słupach bocznych rdzenia kręgowego od poziomu T1 do L2 (neurony przedzwojowe). Wychodząc przez korzenie przednie, biegną razem z włóknami somatycznymi odśrodkowymi od ciał neuronów ruchowych rogów przednich rdzenia i włączają się w nerw rdzeniowy. Na poziomie odejścia gałęzi przedniej i tylnej włókna te opuszczają korzeń i poprzez gałęzie łączące białe (nazwa pochodzi od dużych ilości mieliny w ich osłonkach) wchodzą do zwojów położonych metamerycznie na przednio-bocznych powierzchniach trzonów kręgów lub na poziomie głów żeber. Od tego miejsca istnieją cztery możliwe konfiguracje dalszego przebiegu włókien opuszczających zwoje [3].
- Włókna przedzwojowe mogą przenosić przez synapsy impuls na komórki umieszczone w zwojach i przez gałąź łączącą szarą (nazwa pochodzi z powodu braku mieliny w osłonkach) powędrować z powrotem do nerwu rdzeniowego i dalej do efektorowego narządu na tym samym poziomie metameru T1–L2 (naczynia, mięśnie, okostna, gruczoły skórne).
- Po przełączeniu w zwoju włókno może przechodzić w górę lub w dół do sąsiednich zwojów, a tam przez kolejne przełączenia odnajduje drogę do efektorowych narządów na innych niż pierwotnie zgodnie z metamerem. Stąd np. zwój poziomu Th5 nie tylko przepuści włókno przedzwojowe z metameru Th5 biegnące do swojego efektora w mięśniu sercowym, ale również drogą wstępującą może pobudzić włókna zazwojowe w zwoju szyjnym górnym, stąd poprzez sploty na t. szyjnej (tzw. nerw szyjno-tętniczy wewnętrzny) docierają poprzez splot jamisty do zwoju rzęskowego za gałką oczną i po kolejnym przełączeniu na włókna biegnące do m. rozwieracza źrenicy. Tłumaczy to fizjologiczny odruch poszerzenia źrenicy w sytuacji ostrego niedokrwienia mięśnia sercowego. Oczywiście potrzebne jest wydłużenie zasięgu włókien przedzwojowych, aby mogły zaopatrywać również kończyny górne i dolne, stąd dodatkowe zwoje zlokalizowane nad i pod segmentami rdzenia od T1 do L2, tj. zwój szyjny dolny, środkowy i górny oraz liczne zwoje w części lędźwiowo-krzyżowej położone symetrycznie i zakończone zwojem nieparzystym na poziomie kości guzicznej. Taka budowa zapewnia włóknom z poziomu Th6 a nawet Th7 „wspinaczkę” na poziom zwojów szyjnego dolnego i środkowego i po przełączeniu w nich na włókna zazwojowe zapewnia autonomiczne unerwienie kończyn górnych drogą somatycznych nerwów splotu ramiennego. Ma to doniosłe znaczenie kliniczne, gdyż nader często w sytuacji ostrych zespołów bólowych kończyny górnej, gdzie trudno jest wykorzystać pełną gamę manualnych technik stawowych, oddziaływanie odruchowe poprzez pracę z segmentami odcinka Th i przejścia C/Th może przesądzić o powodzeniu terapii. Podobnie włókna od poziomu Th10, 11 mogą sięgać daleko w dół, wchodząc do somatycznego splotu lędźwiowego i krzyżowego. Zwoje ułożone symetrycznie po obu stronach kręgosłupa i włókna je łączące tworzą pień współczulny. Włókna łączące białe są położone tylko na poziomie od Th1 do L2, natomiast włókna łączące szare towarzyszą wszystkim nerwom rdzeniowym. Włókna wychodzące z segmentów Th1 do Th5 wybierają drogę wstępującą, zaopatrując narządy głowy, szyi, klatki piersiowej. Włókna Th6, 7 mają w tym względzie pewną swobodę – mogą podążać w górę lub w dół do sąsiednich zwojów, od tego poziomu włókna przebiegają już tylko w dół, tworząc zaopatrzenie dla narządów brzucha, miednicy i kończyn dolnych. Zaopatrzenie narządów głowy pochodzi głównie z segmentów Th1 i Th2, a przebiega jak już wspomniano powyżej drogą odgałęzień oplatających naczynia szyjne po opuszczeniu zwoju szyjnego górnego.
- Trzecią możliwością drogi włókien współczulnych po opuszczeniu zwoju jest wyjście ich w formie samodzielnych wypustek tzw. nerwów trzewnych, które zawierają włókna postsynaptyczne z segmentów Th1–5 przełączone w zwojach przykręgowych i tworzą nerwy trzewne sercowo-płucne i przełykowe stanowiące odpowiednie sploty w bezpośrednim sąsiedztwie efektorowego narządu (splot sercowy, płucny, przełykowy).
- Czwartą możliwością przebiegu włókien za zwojem przykręgowym są włókna tranzytowe przechodzące przez zwój przykręgowy bez przełączania i biegnące do zwojów w pobliżu narządu efektorowego (Th6–L2) jako nerwy trzewne brzuszno-miedniczne. Wyróżnia się tu nerw trzewny wielki (Th6–9) idący do zwoju trzewnego, nerw trzewny mały (Th10–11) idący do zwoju krezkowego górnego i nerw trzewny mniejszy (Th12) idący do zwoju aortalno-nerkowego, a także nerwy trzewne lędźwiowe do zwoju krezkowego dolnego i krzyżowe do zwoju podbrzusznego dolnego. Dopiero stąd jako włókna zazwojowe w formie samodzielnych pęczków lub powiązanych ze sobą w sploty tworzą zaopatrzenie narządów wewnętrznych.
Zwoje dla narządów jamy brzusznej i miednicy położone są w sąsiedztwie dużych naczyń, tworząc zwój trzewny, aortalno-nerkowy, krezkowy górny i dolny oraz zwoje podbrzuszny górny i dolny w obrębie miednicy mniejszej. Rozgałęzienia i liczne połączenia tych nerwów, przyjmując przy tym nerwy układu przywspółczulnego, tworzą sploty najczęściej oplatające duże naczynia i ich torem odnajdują drogę do narządu efektorowego (żołądek, wątroba, jelito). Ma to praktyczne znaczenie dla terapii, gdyż znając przebieg dużych naczyń, można według nich dostosować wektor mobilizacji splotów autonomicznych.
Włókna przedzwojowe, których punktem wyjścia są komórki w mózgu lub rdzeniu, są zmielinizowane – szybko przewodzące, dodatkowo włókna przedzwojowe od rdzenia do zwoju przykręgowego mają zaledwie 2–3 cm, natomiast włókna pozazwojowe są niezmielinizowane, a więc znacznie wolniej przewodzą bodźce i osiągają długość nawet kilkunastu centymetrów. We włóknach przedzwojowych zarówno układu współczulnego, jak i przywspółczulnego, neurotransmiterem jest acetylocholina (podobnie jak w układzie somatycznym), natomiast we włóknach pozazwojowych z wyjątkami gruczołów potowych (acetylocholinowe) neurotransmiterem bodźca na narząd docelowy dla włókien współczulnych jest noradrenalina, a dla włókien przywspółczulnych – acetylocholina. Szczególnym rodzajem narządu efektorowego jest dla włókien współczulnych przedzwojowych rdzeń nadnerczy, do którego docierają przedzwojowe włókna przechodzące tranzytem przez zwój trzewny, a ...
